
Excerpted from Garden of Marvels: How We Discovered that Flowers Have Sex, Leaves Eat Air, and Other Secrets of Plants by Ruth Kassinger, out now from William Morrow.
Why flowers, anyhow? Plants began to conquer the land more than 400 million years ago and ruled over it for more than 250 million years without producing a single blossom. Why should they have? Flowers are expensive. Sepals, petals, pigments for color, organic compounds for scent: Creating those fancy clothes and complex perfumes takes a lot of stored energy. Instead of manufacturing flowers, a plant could have used those carbohydrates to make more seeds or grow taller, both proven strategies in the competition for survival. Besides, there seems to be nothing in gymnosperms (nonflowering plants) that corresponds to flowers. Angiosperms (flowering plants) seem to have arisen out of nothing, sui generis. Nonetheless, blossoms—from the oak’s minuscule brown nubs to the green spikelets of rice to the multipetaled splendor of the rose—appear on at least 75 percent of all the world’s plant species.
The why and how of angiosperms, Darwin wrote in 1879, are “an abominable mystery.” The mystery still has not been fully solved. Part of the difficulty is that flowers have always been fragile and when they die, they fall apart into easily scattered and perishable pieces. The fossil record of early flowers is therefore exceedingly scant. In recent years, however, evolutionary botanists have come to think the living Amborella trichopoda will help solve the puzzle.
You will never find anyone who will sell you a bouquet of amborella. For one, the cream-colored, dime-sized flowers on this knee-high shrub are not much to look at, and will set no lover’s heart aflutter. If you should want to buy an amborella plant (say, you want to write about it), you’ll also be out of luck. The shrub is rare, and grows naturally only in the cloud forests of the island of New Caledonia in the South Pacific. Only a few American conservatories cultivate amborella, which is notoriously difficult to sustain and which flowers unpredictably in captivity. Nonetheless, unprepossessing and persnickety as it is, the plant has attracted a great deal of scrutiny recently. It is likely the closest living relative to the first flowering plant, and according to Harvard professor William Friedman, “a critical missing link between angiosperms and gymnosperms.”
About 140 million years ago, at the beginning of the Cretaceous era, a new type of plant evolved from one of the seed-bearing, nonflowering gymnosperms—mostly conifers—that covered much of the landscape. This first angiosperm was the founder of a new line, the Amborellales. Sometime later a sister angiosperm line emerged, the Nymphaeales, which evolved to become modern water lilies. A third line emerged. These, the Austrobaileyales, evolved rapidly and diversified profusely to become almost all of today’s 250,000 to 400,000 (depending on who’s counting) flowering species, from cucumbers to pansies to elms. The Amborellales, on the other hand, are today represented by a single species, Amborella trichopoda, the modern flowering plant least changed from its gymnosperm ancestor. It is a living relic from the age of the dinosaurs.
So, what does amborella have to say about why angiosperms evolved and conquered? Talking about angiosperms is talking about sex. Gymnosperms are dioecious. For example, individual gingko trees, which are one of the few nonconiferous gymnosperms, bear either male or female organs. Among conifers, male and female cones can be on separate individual trees or, more often, segregated on either the lower or upper branches of a single tree. (The male cones, which have pollen sacs on the upper edge of their scales, are smaller and drop soon after they release their pollen. Female cones grow larger after pollination, taking anywhere from a few months to two years before falling.) On the other hand, most angiosperms are hermaphrodites, with stamens and carpels inside the same the flower.
Amborella is in a confused state when it comes to sex, as if transitioning from gymnosperm to angiosperm anatomy. It has separate male and female organs on the same plant, like a conifer. The male flowers have stamens only, but they don’t look like modern stamens—that is, with filaments topped with two pollen-bearing sacs. Instead, the two pollen sacs are carried on the edge of flat and broad petals that look very much like the scales in male conifer cones. But they also have flowers that look like hermaphrodites, with both carpels and stamens. These stamens, however, are sterile, making them staminodes.
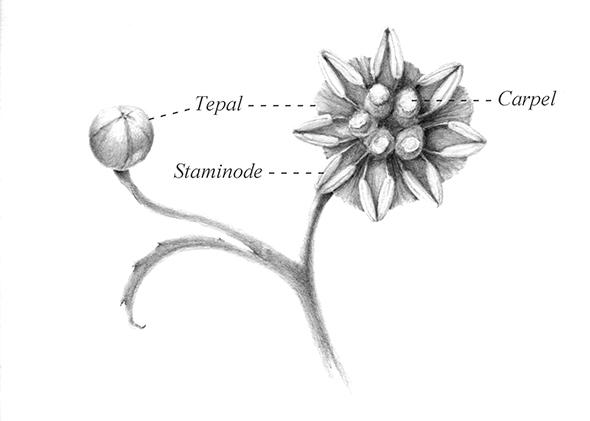
Illustration by Eva-Maria Ruhl.
How did these other flowers (which function as female flowers) manage to acquire both carpels and nonfunctional stamens? The debate is far from over, but it seems the first and critical step was a “whole genome duplication” in the common ancestor of all flowering plants. A gymnosperm (like a human) is diploid, which means it has one copy of its genome in the nucleus of all of its cells. (The term diploid indicates that the genome is a set of paired chromosomes, one from each of its two parents.) After the whole genome duplication, the mutated plant and its tetraploid descendants had two copies of its genome in the nucleus of its cells. These plants became instant winners in the survival game. They now had spare genes for experimenting, spare genes that might mutate freely without endangering the viability of the individual. If a mutation in a spare gene conferred an advantage, the mutation could be passed on. If the spare gene mutated in an unproductive way, it would be lost.
Today most flowering species have more than one copy of their genomes, and thousands have a dozen or more copies. Plants with multiple copies are called polyploids, and tend to be larger, have more complex flowers, and bear bigger fruits than their genomically less endowed relatives. Humans have shaped plant evolution by selecting polyploid plants for cultivation. Modern garden strawberries, for example, are octoploids, and some have berries so large they need to be cut in quarters to be eaten, while their wild relatives are diploid or at most tetraploid and have berries the size of a pea. Plant breeders often propagate garden flowers with doubled genomes to produce double the usual number of petals. In the era just before the amborella emerged, however, a doubled genome simply meant a new plasticity for basic forms and functions. That plasticity was critical to evolving leaves into colorful petals and into sepals, the leaves that protect unopened buds. Gene redundancy also was a factor in developing male and female organs in the same flower.
Gender in gymnosperms of the era must have already been nuanced and mutable. Only four groups of gymnosperms are still living today, but some of their members have interesting sexual variations. Some gymnosperms change genders as they mature. Male gingkoes occasionally metamorphose into females, much to the distress of city residents. (Gingkoes are popular urban street trees—they’re so tough, some survived the nuclear blast in Hiroshima—but female seeds are terribly messy and their odor has been compared to that of vomit or rancid butter.) The Mediterranean cypress starts off female but later becomes male. Many conifers have their male cones on the lower branches and female cones on the upper, but a spruce and several pine varieties are sexually upside-down. So, when the gymnosperm genome first doubled and was chock-full of extra and as-yet-unassigned genes, gender would have been particularly malleable. Amborella, with its flowers that are unisexual in function but hermaphrodite in appearance, is not a shocking experimental result. And amborella is known to go transgender, too.
One of the most important innovations of amborella and later angiosperms was to enclose its seeds in an ovary. No gymnosperm seed has an ovary: Their fertilized ovules are “naked,” covered only in a single, thin protective layer. An angiosperm’s ovary protects its embryo from desiccation and physical harm. Some species’ ovaries enlarge significantly to become edible fruits. If you’re a plant and you want to have your seeds spread widely, fruits are the way to go. Animals transport the fruits inside their digestive tracts, and then deposit the embryo far from the parent, along with a nice pile of nitrogen-rich manure. Botanically speaking, grains and nuts are also fruits. It’s just that their ovaries are dry and tough.
So where did the ovary come from? Gene duplication again. The ovary is the bottommost part of the carpel, and the carpel, genetic analysis shows, is a repurposed leaf, which by folding and fusing its edges together came to form a second, impermeable covering over the seed. In fact, the amborella carpel has a visible seam where the leaf has not quite fully fused. An edible fruit is not the only transportation method that angiosperms developed for their seeds. Thanks to those duplicated genomes, external seed structures, such as burrs and hooks, plumes, and sticky liquids, have evolved to send seeds far down the road. Wait a few million years, and angiosperm seeds will be hailing cabs.
Excerpted from Garden of Marvels: How We Discovered that Flowers Have Sex, Leaves Eat Air, and Other Secrets of Plants by Ruth Kassinger, out now from William Morrow.